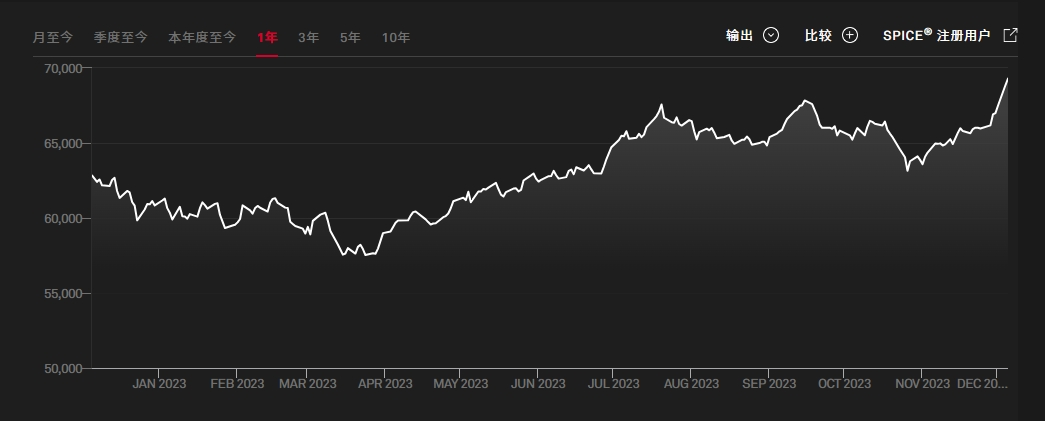
起源演化
侏罗纪时期
侏罗纪-白垩纪分界线的化石表明,水蚤类起源超过1.45亿年,Ctenodaphnia亚属和水蚤亚属(Daphnia)可能起源于2亿年前的侏罗纪,也就是盘古大陆分裂成北方的劳亚大陆和南方的冈瓦纳大陆的时期。水蚤属的异源物种占到物种形成过程的42%(30%是由于洲际分裂,12%是由于大陆内分离),并且欧洲和北美的对应物种显示出高度的序列分化,从而表明水蚤是在2~3亿年之间分裂。
白垩纪时期
水蚤在7000万年时已经多样化,再加上更新世(距今260万年至1万年)的反复冰期而循环塑造了水蚤当前的地理分布,水蚤的地理分布显示出地理上的二分法,Ctenodaphnia亚属多出现在南半球,而水蚤亚属(Daphnia)多出现在北半球。比如末次冰期期间向高北极避难所转移的冰川导致了蚤状水蚤(Daphnia pulex)向格陵兰岛,冰岛,斯瓦尔巴群岛北部部分地区的迁移和定居;在古北区(包括欧洲,北回归线以北的非洲与阿拉伯半岛中北部以及喜马拉雅山与秦岭山脉以北的亚洲大陆),发现长刺水蚤(daphnia longispina)起源于500到700万年前,其原因是由于当时多个冰川避难所的范围扩大而让它们在此定居;隆线水蚤(Daphnia carinata)在澳大利亚雪山高山冰湖的有限分布也归因于冰川作用。
演化加速
栖息地的变化加速了水蚤的进化速度,塑造了不同生活史特征、形态特征、能量代谢等生物功能和行为。当潜在的杂交物种由于无法有效利用彼此的栖息地而在繁殖期间没有遇到彼此时就会发生栖息地隔离。例如栖息地的不同会导致水蚤的反捕食行为也不同,栖息在小湖泊的Daphnia parvula被无脊椎动物捕食的风险较低,而栖息在相对更大的栖息地的Daphnia retrocurve则是更有可能被捕食,所以它表现出突出的头盔作为自己的防御手段。同时,水蚤也会因为栖息地的不同而表现出不同的滤网尺寸和生态偏好。除了栖息地外,不同种类繁殖时间的差异、行为的不同以及杂交种存活率低等方面都对水蚤的基因流动产生了或多或少的影响。
杂交在自然界中很常见,水蚤属的近亲物种也会经常杂交,并有可能产生新物种,例如在更新世冰期的时候,Daphnia pulicaria与Daphnia tenebrosa或Daphnia melanica杂交而产生了一个新物种Daphnia midendorffiana。同时,水蚤的种间杂交也会产生具有高适应潜力的多倍体从而更好的适应环境。
命名与分类
1785年,丹麦动物学家穆勒(O.F.Müller)首次对水蚤属进行了正式描述,当时这一属下已经有38种水蚤被确认。而发展至今,随着动物研究人员的不断观察,有越来越多的水蚤属的物种被他们发现并命名。它们都被记录在物种分类网站中,但由于调查方向的不同,各个分类网站对水蚤属的分类也不尽相同。根据全球生物物种名录(COL)的记录,截至2013年,水蚤属在全世界发现并命名了3亚属,200种。而根据世界海洋物种目录(WoRMS)的记录,截至2023年,水蚤属在全世界发现并命名了3亚属,332种。本百科根据全球生物物种名录(COL)的分类展开,得知水蚤属的3个亚属分别为Australodaphnia亚属、Ctenodaphnia亚属、水蚤亚属(Daphnia)。具体分类情况及代表物种如下表:
亚属学名 | 亚属下物种数量 | 物种 | 代表物种图片 |
Australodaphnia亚属 | 1个 | Australodaphnia occidentalis | 无 |
Ctenodaphnia亚属 | 46个 | 大型水蚤(Daphnia magna)、隆线水蚤(Daphnia carinata)、Ctenodaphnia barbata、Ctenodaphnia bolivari、Ctenodaphnia brooksi等 | 大型水蚤 
隆线水蚤 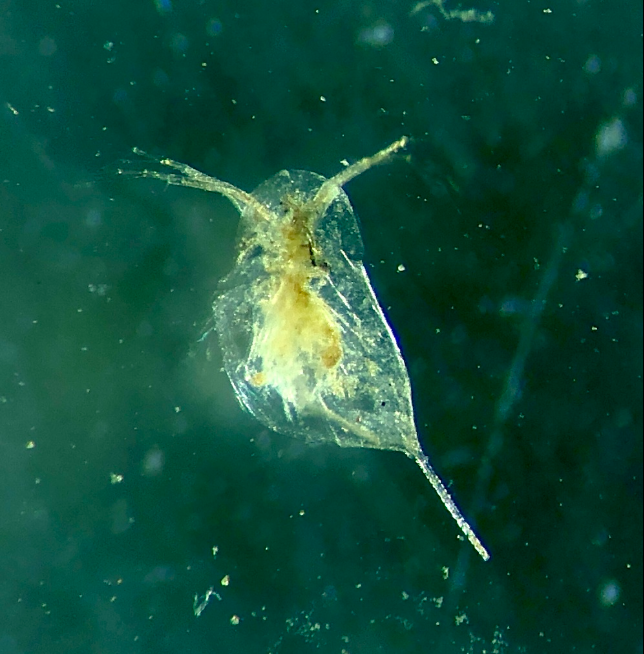
|
水蚤亚属(Daphnia) | 72个 | 僧帽水蚤(Daphnia cucullata)、透明水蚤(Daphnia hyalina)、长刺水蚤(Daphnia longispina)、蚤状水蚤(Daphnia pulex)等 | 僧帽水蚤 
透明水蚤 
|
形态特征
水蚤成体体长一般为1-5毫米,大型水蚤体长可达6毫米。水蚤体呈卵圆形或椭圆形,比较侧扁,有扁平的叶状腿。它们的身体被一层未钙化的壳所包围,称为甲壳。它有两层壁,血淋巴在两层壁之间流动,这是体腔的一部分。甲壳主要由几丁质构成,这是一种多糖。甲壳背面具有脊棱。后端延伸而成长的壳刺。后端部分以及壳刺的沿缘均被有小棘。壳面有棱形和多角形网纹。头部与躯干部的界限不是很清楚,但负有冬卵的雌体可明显地分为头与躯干两部分。作为双甲总目(Diplostraca)下的物种,水蚤有多达10对附肢,分别是(从前到后):触角、触角(第二个触角,用于游泳),上颌骨和下颚,其次是躯干上的5个肢体。四肢形成进食和呼吸的器官。腹部的末端有一对爪子。
水蚤雌体通常无颈沟。吻明显,大多尖。一般都有单眼。第一触角短小,部分或几乎全被吻部掩盖,不能活动。绝大多数种类的第二触角共有9根游泳刚毛,但小栉只有8根。腹部被侧有3~4个发达的腹突。靠近前部的腹突特别长,呈舌状,伸向前方。后腹部细长,由前后逐渐收削。卵鞍近乎巨形或三角形,内储2个冬卵。
水蚤雄体较小,壳瓣背缘平直。前腹角凸出,列生较长的刚毛。吻无或十分短钝。第一触角长大,能活动,通常具有粗长的鞭毛。第一胸肢有钩与鞭毛。腹突常退化。
水蚤属下的物种还会呈现出不同的体色。例如隆线水蚤是浅黄或稍带红色,透明水蚤是淡灰色或浅绿色,长刺水蚤是黄、红、棕、浅绿或深灰色,大型水蚤是黄色或淡红色,蚤状水蚤是半透明,带浅黄棕色或淡绿色。
分布栖息
分布范围
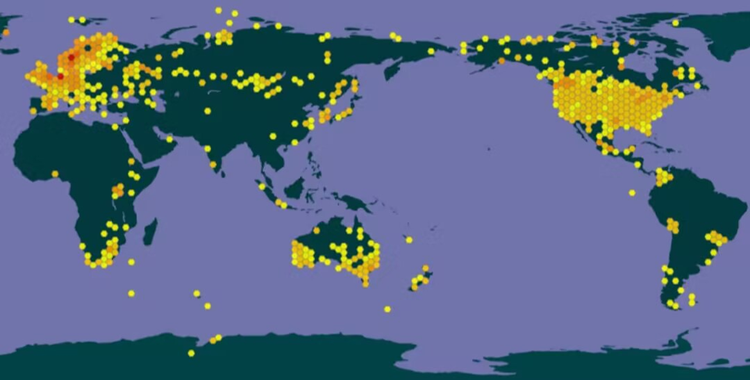
水蚤分布于世界各地,尤以温带最为普遍。在北美、欧洲和澳大利亚等处有大量分布。从下面的水蚤分布图也可以看出水蚤在非洲南部、亚洲西部中部及北部、南美洲北部和西南东南部有少量分布。
栖息环境
水蚤主要栖息在湖泊、池塘和溪流中,水蚤属下的不同物种的栖息环境也有所不同,例如隆线水蚤(Daphnia carinata)喜居于富营养型小水域中,但也出现于湖泊、水库以及江河中,有时也在咸水中生存;透明水蚤(Daphnia hyalina)主要栖息于较大的贫营养型湖泊中,是敞水区典型的浮游种类;长刺水蚤(Daphnia longispina)居于水草繁茂的富营养型小水域中,例如池塘、水坑、间歇性积水以及小型湖泊等;蚤状水蚤(Daphnia pulex)在北纬和中纬地带,是水潭、水坑、池塘以及小河等小型水域中的优势种类,尤其在富营养型水域中,分布特别普遍。在南纬地带,出现于湖泊或水库等敞水区。水蚤栖息地的水质变化很大。pH值在6.5至9.5之间对大多数物种来说是可以接受的,最佳pH值在7.2至8.5之间。盐度通常应低于海水的5%(约每升1.5克海盐),但有些物种可以忍受更高的盐度,例如大型水蚤(Daphnia magna)可以在高达20%的海水中找到。
习性
觅食行为
水蚤喜欢以藻类为食,在水生系统中,碳、磷和氮等元素的比例用于表征藻类的食物质量。当食物质量呈现不同空间梯度时,水蚤迅速找到(即<5-10分钟)高食物质量的空间区域,并改变其觅食行为以留在这些区域。幼体和成体水蚤都能够在大致相同的时间尺度上快速定位食物质量高的区域并表现出相似的行为,它们的觅食路径会以食物质量高的地区为中心,个体可以在快速的行为时间尺度上调整它们的摄入率。相反,暴露于食物质量和数量的均匀环境中的水蚤则会随机觅食。
水蚤是一种非选择性滤食性动物,食物通过过滤装置收集,而腿使一些水从前流向后,并通过刚毛将颗粒移动到肠道。例如大型水蚤(Daphnia magna),它们的主要取食策略是过滤悬浮颗粒提取营养物质,如绿藻。由于藻类已被发现在塑料基质上生长并形成生物膜。因此,它们会摄入微塑料珠,然后从表面消化藻类。一旦藻类被消化,微塑料珠就会被排出体外,从而为新珠及其相关藻类的摄入创造空间。
防御行为
水蚤在应对捕食者的捕食时,会表现出诱导防御行为,而诱导防御也会推动水蚤的生活史、行为和形态特征的进化。在水蚤中,这些性状表现出相当大的表型可塑性(即同一生物应对不同的环境影响而作出不同的改变),这使得水蚤能够适应环境的变化。鱼类释放的利他素会刺激它们的生长。它们会加速发育,以便在更早的年龄和更小的体型成熟,进而它们能在相同的时间内获得更多的成体。它们还会在捕食者存在的情况下也会开始更早地繁殖,并且产生额外的幼体。
同时,水蚤对于不同的鱼类体型还会产生不同的反应。例如,体型相对更大的太阳鱼( Lepomis gibbosus)比食蚊鱼(Gambusia affinis)更显著地提高了的长刺水蚤(Daphnia longispina)繁殖能力。这是水蚤对不同尺寸鱼类的选择性捕食而导致的大量死亡做出适应性的“积极”反应。为了应对捕食者的捕食,某些水蚤也会发生形态特征的变化,例如Daphnia atkinsonspecies在欧洲鲎虫(Triops cancriformis)释放的化学物质的刺激下产生了“刺冠”。在蚤状水蚤(Daphnia pulex)和长刺水蚤(Daphnia longispina)中都发现了颈齿形成等形态防御,但它们通常在蚤状水蚤(Daphnia pulex)中更为明显。
游泳行为
水蚤在水中的游泳行为包括水平、直线、垂直、下沉、跳跃等等。雄性水蚤和雌性水蚤的游泳模式不同,雄性水蚤游的更快,而且是垂直的。在较小的容器中,水蚤属下无论什么物种,游得都更慢,下沉速度也较慢,并且倾向于沿直线移动。水蚤的游泳行为会受到光照、食物和捕食者的影响。一般来说,在较高的食物浓度下水蚤往往表现出较快的速度和较高的下沉率,但对转弯角度没有一致的影响。在较亮的光下水蚤往往表现出较慢的速度和较低的下沉率,并且具有较小的转向角,当灯打开时,水蚤以快速向下游动的方式向下移动,而不是下沉或适度游动;当灯关闭时,水蚤通过快速向上游动而不是适度向上游动。水蚤的普通游泳接近水平,快速游泳接近垂直。
鱼的气味以两种方式影响水蚤。鱼腥味的存在增加了在光线变化后表现出快速游动的种群的比例,并且在有鱼腥味的情况下,下沉行为往往更少。然而,也有一些例外。例如,大型水蚤(Daphnia magna)在高食物和高光照的条件下游得最快。
游得更快的个体可能更有效地觅食,但也可能更容易受到捕食者的攻击。但快速垂直游动比下沉或适度向下游动导致的死亡率更低。快速游动的水蚤几乎肯定比缓慢游动的水蚤更引人注目,然而,快速游动的水蚤在明亮的上游水域待的时间较少,从而减少被视觉捕食者发现的时间。而在昏暗的光线下向上快速游动,水蚤被鱼捕食的风险也很小,因为鱼很难看到水蚤。
对于水蚤来说,跳跃是它们的一种常见游泳行为。水蚤每天可以游几米,而且成年水蚤通常比幼年水蚤游得更远。不同体型和不同种类的水蚤会表现出相似的行为以最大化的提高自己的摄食率和降低死亡率。同时,水蚤的游泳行为还会受到悬浮酵母浓度的影响。在高浓度酵母环境下,水蚤会减少它们在垂直方向上的游动,转而沿着直线游动。
迁移行为
水蚤分布的普遍性是由于它们能够耐受一系列动态的环境条件,特别是温度。水蚤每天在水体内迁移以避免捕食,这种行为称为昼夜垂直迁移(DVM)。水蚤的经典DVM行为是在白天迁移到更深,更暗和更冷的水域以避免视觉捕食者,并在夜间迁移到较温暖的表层水域,以便在被捕食风险降低时进食。DVM受到距离和水中铜镍水平的影响,添加铜镍中的任何一种都会抑制水蚤在光照存在下对鱼类利他素的反应,从而使这些迁徙的水蚤面临更大的捕食风险。
聚集与分散
浮游动物中,水蚤的聚集行为是一种避免捕食的行为。该种行为具有两个作用,分别是混淆捕食者的视觉和减少个体被吃掉的几率。但聚集行为也会有一定的坏处,就是会让聚集区域的食物浓度快速降低,从而会使水蚤因为缺乏食物而分散,所以在食物过量的情况下水蚤往往更容易聚集。以挪威米拉瓦特湖沿岸的长刺水蚤(Daphnia longispina)为例,它们会在沿岸发生聚集,而且夜间群体密度较低,约为白天的十分之一。群体在夜间的分散表明了一种躲避行为,以应对视觉上或白天活动的捕食者。

生长繁殖
交配方式
水蚤在有性繁殖期间会表现出群体行为,雌体可能会被多个雄体受精。在群体中,雄体会通过多种方式来定位雌体,例如雄体会在水中快速垂直游动以提高跟雌体的相遇率,这种行为被称为“扫描”。除“扫描”外,雄体还能通过机械接收特定的游泳和过滤行为产生的流体干扰来定位雌体。雄体对雌体的生殖阶段还有某种形式的认识,它们会更倾向于追求具有性接受能力的雌体而不是那些没有的。同时,雌体对雄体也会有一定的选择,一旦雄体与雌体开始结合,位于雌体卵巢中的糖蛋白可能会为配偶识别提供区分信号,在结合过程中中,雄体分离之前大约15秒就会发生交配,而对于其它种类的甲壳类动物,交配可以持续几分钟到几天。除同种物种间的交配外,水蚤属中的血缘相近不同物种有时会相互杂交,但与亲本相比,杂交种的孵化率、存活率和对环境的适应能力都较低。
繁殖产卵
水蚤有孤雌生殖和有性生殖两种生殖方式。当环境适合水蚤的生活时,就会发生孤雌生殖。水蚤的个体均为雌性,但当环境质量较差时,水蚤会产生雄性进行有性繁殖。水蚤属中的许多水蚤都是以孤雌生殖和有性生殖相结合的多周期生殖为主要繁殖方式,比如隆线水蚤(Daphnia carinata)、透明水蚤(Daphnia hyalina)、蚤状水蚤(Daphnia pulex)、长刺水蚤(Daphnia longispina)、大型水蚤(Daphnia magna),这种生殖方式也让它们能更好的适应环境。

水蚤的产卵间隔因多种因素而异,在有利条件下通常需要约3-4日。卵母细胞在成熟过程中会积累卵黄颗粒和油滴,待到成为完全长大的卵子大约需要 60 小时,在这个过程中卵会在育雏室中发育成新生儿,并在雌体蜕皮后13分钟被产出。大型水蚤每只成年雌体每龄产6至50只幼虫(大小约为1毫米),龄期为2 - 3天。水蚤的产卵量还会因温度的变化而变化。水蚤还会产生一种休眠胚胎,它被包裹在一种被称为卵鞍的坚硬结构中被提供机械保护,并抵抗干燥和冷冻。卵鞍可以通过矢量(如水流、风、动物)在不同地点之间传播,也可以通过沉积物中的沉积和潜在的孵化来穿越时间。包裹在卵鞍中的胚胎可以在未受干扰的沉积物中休眠数十年或数百年。并能在适应的条件下自行孵化。
水蚤的繁殖会受到多种因素的影响,其中最为典型的就是食物和捕食者对水蚤繁殖的影响。例如,以缺氮藻类为食的水蚤能将更多的脂质分配给卵,其后代在饥饿条件下寿命更长。与高食物水平相比,相同体长的个体在低食物水平下的生殖投资相对较少,在低食物水平下生长的水蚤的育雏室明显小于在高食物水平下生长的相同体长的水蚤,并且在考虑育雏室体积与卵体积的比例的情况下,相对较小的水蚤所产的卵相对较大从而有更多的母体储备,可以在早期新生儿开始有效地摄入和消化藻类食物之前提高存活率。现在有相当多的证据表明,捕食者释放的化学物质会影响其潜在浮游动物的繁殖。高密度浮游动物释放的化学物质也会抑制它们自己和其它浮游动物物种的摄食和繁殖。大仰蝽属(Notonecta)产生的某种水溶性化学因子引起了Daphnia galeata胚胎后期发育过程中能量分配模式的改变,使其分配给增长的那部分能量减少了,而分配给再生产的能量增加了。
生长发育
蜕皮是水蚤生长的必经之路,水蚤只有在蜕皮后,当被皮仍然柔软时,才会立即生长,因此它们的生长是不连续的,一只水蚤一生中要蜕10-18次皮。在蜕皮的过程中,水蚤会流失钙、磷、碳等多种元素。水蚤的生长也同样会受到捕食者和食物的影响。例如鱼类释放的利他素会加速水蚤的生长,在高食物浓度下,缺钙会让水蚤的碳积累受到限制从而使其体重增加减少。大型水蚤在离开育雏室后6至8天内成熟,在实验室条件下水蚤可以存活两个月以上。[43]大型水蚤在20摄氏度时的寿命相对较短(约8周)。
人工饲养
水蚤的人工饲养活动包括工具材料的制备、灭菌、水蚤幼苗的供应、培养基的制作、培养、收获和水质监测。
饲养条件
水蚤的生长速率受食物来源、种类和环境等因素的影响。养殖水蚤的食物来源很多,如有机肥,尤其是鸡粪、牛粪、羊粪等。水中的粪便会产生细菌、碎屑和其他物质,它们会成为水蚤的食物。氨是水蚤培养的关键,是动物粪便的主要成分。氨以尿酸的形式存在于家禽粪便中,以尿素的形式存在于哺乳动物粪便中。也有实验表明,使用鹌鹑粪养殖的水蚤生长速率和繁殖率都高于用鸡粪养殖的水蚤。
饲养方法
以实验室饲养水蚤的方法为例,实验室规模培养中使用的容器由透明玻璃水族箱制成,其注水量为水深达15厘米或容量达到31.5升。而后对充满水的水族箱进行曝气。
在饲养后第1天至第4天的适应阶段,水蚤生长相对缓慢。水蚤个体数量增加后第4天进入指数增长阶段并且在5-7天内增加最多,其原因是第4天之后水蚤种群增加是因为在环境条件或培养基可育性达到一定程度的情况下,发生了产生水蚤个体的无性繁殖(孤雌生殖)过程。在第7~10天进入稳定期或滞后期,由于培养液中营养物质的利用率不能满足培养皿中水蚤的需要,水蚤的种群增长率下降。水蚤在培养基中的最优种群生长可达1500-5000个。
在水蚤幼苗放养后第4天可进行部分收获。一次部分收获小于培养基总积的20%。可在上午和下午进行收获。全部采集可在两种情况下进行,即达到最高密度使水蚤数量降低时,必须进行采收。水蚤的采收有两种不同的方法,即部分采收法和全部采收法。收获的水蚤通常用作幼年鱼类饲料,如鲶鱼、罗非鱼和鲤鱼。
饲养照料
温度、酸碱度和溶解氧,对水蚤的养殖至关重要。酸碱度影响浮游动物微甲壳类动物卵的寿命,水蚤培养的最佳pH值在7~8.6之间。中性或碱性有利于大型水蚤的生长。pH值还决定了氨对培养基的毒性。氨对所有生物都有很大的毒性,但在酸性条件下,氨的毒性会降低,因此需要对水蚤培养进行pH监测。此外,还要通过曝气控制培养物的DO在水蚤的耐受范围内,氧气也使微生物有机会进行代谢,分解介质中的有机物,使水质良好。养殖水蚤在20℃温度下表现良好。必须控制好温度。
此外,大型水蚤(Daphnia magna)的养殖还会受到水的硬度的影响,随着养殖水硬度的增加,其幼体的数量和产卵量都在增加。此外,性成熟的时间随着水硬度的增加而稳步减少。在碳酸钙的平均硬度为350毫克每升的培养水中饲喂绿藻、硒酸盐以及鳟鱼饲料和苜蓿的混合物的组合,大型水蚤的生产力最高。
物种保护
物种现状
20世纪末,在《世界自然保护联盟(IUCN)濒危物种红色名录》的统计数据中,现存于南非的水蚤属物种Daphnia coronata,和现存于澳大利亚的水蚤属物种Daphnia occidentalis、Daphnia jollyi、Daphnia nivalis。这四种水蚤所栖息的环境要么范围变小要么质量下降,这导致了它们成体数量的持续下降。除此四种之外的水蚤属节肢动物种群资源尚不存在较大的威胁。
保护级别
截至1996年,水蚤属物种有4种被记录在《世界自然保护联盟(IUCN)濒危物种红色名录》中,具体保护情况如下表:
物种名 | 濒危级别 | 评估时间 |
Daphnia coronata | 易危(VU) | 1996年 |
Daphnia occidentalis | 易危(VU) | 1996年 |
Daphnia jollyi | 易危(VU) | 1996年 |
Daphnia nivalis | 易危(VU) | 1996年 |
致危因素
气候变化、人为影响和外来物种入侵都对水蚤的生存产生了极大的影响。例如,由于气候变化,地表水温度继续升高,甚至像北美五大湖这样的大型透镜系统也预计会变得更加多变,导致垂直混合的变化以及湖泊热分层的程度上升,湖泊分层的变化会影响捕食者-猎物动态,因此可能会影响水蚤的分布和适应性,使水蚤受到温度的严重影响而导致湖泊内水蚤种内竞争加剧,全球水蚤多样性下降。加拿大许多湖泊钙含量的下降也导致了水蚤群落的生产力和结构。人为投入水中的镍、铜等元素会影响水蚤对鱼类利他素的反应从而面临更大的捕食风险。位于美国麦迪逊市的门多塔湖因为外来物种的入侵而让当地的水蚤种群数量大幅降低。
主要价值
经济价值
水蚤属下的部分物种具有一定的经济价值,例如蚤状水蚤(Daphnia pulex)参与了许多维持我们消费或商业使用的鱼类所必需的食物链,在研究捕食者猎物关系时,水蚤被选为“标准猎物”,如鲈鱼、棘鱼、鲦鱼和年轻的红鲑鱼。它们也是鳟鱼和许多其它流行鱼类所依赖的动物的主要食物供应。大型水蚤(Daphnia magna)由于易于养殖,通常喂给在水族箱中饲养的鱼。
科研价值
水蚤已被用作生态学、生态毒理学和进化的模式生物,并且是研究最多的双甲总目(Diplostraca)之一,随着人类在淡水系统中活动的增加,淡水系统的生物多样性正在急剧下降,而对于水蚤物种形成的研究能方便人们了解人类活动对淡水系统多样性造成的影响并作出相应措施。
蚤状水蚤(Daphnia pulex)经常被科学家用于实验。它们体积小,价格便宜,并且很容易在实验室环境中保持活力。它们几乎透明的外壳使它们的内部功能更容易研究,并且它们非常容易受到温度、食物供应或环境中溶解氧含量变化的影响。水族馆所有者经常使用水蚤作为鱼类的食物来源和清除水中的碎片。
生态价值
水蚤以藻类为食,也是小型鱼类的主要食物,在生态环境中提供重要的生态系统服务,如养分循环、食物网动态和生物指标,有着将初级生产者的生物量转移到食物链上的核心作用。几乎所有淡水生态系统都依赖于蚤状水蚤(Daphnia pulex)将浮游植物和腐烂物质转化为更可用形式的能力。
水蚤还有有净化和指示水质的作用,如果生态系统中的水蚤数量大量减少,该生态系统功能就可能被破坏。例如位于美国麦迪逊市的门多塔湖的水蚤因为外来物种长柱尾突溞 (Bythotrephes longimanus)捕食而导致湖水清澈度下降近1米,造成了每年140.2016亿美元的损失。大型水蚤(Daphnia magna)可以通过食用水中可能积聚的藻类和其他碎屑在池塘和湖泊中提供更清洁的水。它也是水质的指示生物,用于水毒性测试和检测各种污染物。
危害
水蚤虽然对生态环境有着十分积极的作用,但它的危害也不容忽视。例如蚤状水蚤(Daphnia pulex)会与水中的其它生物争夺食物和氧气而限制它们的种群规模。虽然它们经常用于鱼缸中以清除含有藻华的水,但由于氧气供应的限制,鱼无法与大量蚤状水蚤(Daphnia pulex)饲养在同一水箱中。