分类争议
带鱼科鱼类的外部形态特征相似,一些分类关键问题存在争议,难以确定某些物种。直到20世纪90年代,有的分类学家还认为中国只有一种带鱼。据《中国生物物种名录》2023版显示,带鱼科隶属于辐鳍鱼纲鲈形目鲭亚目,包括7属,分别是剃刀带鱼属(Assurger)、深海带鱼属(Benthodesmus)、小带鱼属(Eupleurogrammus)、窄颅带鱼属(Evoxymetopon)、沙带鱼属(Lepturacanthus)、狭颅带鱼属(Tentoriceps)、带鱼属(Trichiurus)。2006年,加拿大著名鱼类学家约瑟夫·纳尔逊(Joseph S. Nelson)将原属于带鱼亚目(Trichiuroidei)的带鱼科并入鲭亚目(Scombroidei)。2016年,Nelson将鲭亚目重新确认为鲭形目(Scombriformes)。但截至2023年,《中国生物物种名录》并未接受鲭形目的分类地位,带鱼科仍隶属于鲈形目。
带鱼科有三个亚科,分别是Aphanopodinae亚科,2属;Lepidopodinae亚科,4属;Trichiurinae亚科,4属。截至2023年,国际综合生物分类信息系统(ITIS)和鱼类数据库(FishBase)显示带鱼科共10属46种。另外,捷克BioLib国际生物图书馆记录带鱼科还有一化石类群Anenchelum。
本词条依ITIS所记录的具体属信息如下:
中文名称 | 学名 | 命名者与年份 | 物种数 |
短尾带鱼属 | Aphanopus | Lowe, 1839 | 7 |
棘背带鱼属 | Assurger | Whitley, 1933 | 1 |
深海带鱼属 | Benthodesmus | Goode & Bean, 1882 | 11 |
新月带鱼属 | Demissolinea | Burhanuddin & Iwatsuki, 2003 | 1 |
小带鱼属 | Eupleurogrammus | Gill, 1862 | 2 |
突额带鱼属 | Evoxymetopon | Gill, 1863 | 3 |
鳞带鱼属 | Lepidopus | Goüan, 1770 | 6 |
沙带鱼属 | Lepturacanthus | Fowler, 1905 | 3 |
狭颅带鱼属 | Tentoriceps | Whitley, 1948 | 1 |
带鱼属 | Trichiurus | Linnaeus, 1758 | 11 |
形态特征
带鱼科鱼类身体颇延长,体长最小的是小带鱼属,一般为15cm~50cm;体长最大的是棘背带鱼属的棘背带鱼(Assurger anzac),可达225cm。侧面扁平,整体呈带状,身体颜色为银色至黑色,深海带鱼的体色为黑色或深色,而生活在较浅海域的带鱼体色较浅。体长可达200cm以上。
头长,背面平坦或微凸,或侧扁、高锐突起。吻部长且尖锐。口大,上颌骨被眶前骨覆盖,下颌突出。上下两颌齿强大,尖锐而侧扁。眼睛大,位于侧面。两侧各有一个鼻孔,呈椭圆形或细长形。鳃盖碎裂。鳃耙有刺。梨骨无齿,颚骨具细齿。
鳞退化或消失。侧线1条,连续,伸至尾鳍。背鳍基底甚长,起点始于鳃盖上方,延伸至尾端,硬棘部及软条部连续难区分,硬棘部通常短于软条部,某些种类鳍棘部与鳍条部之间有凹刻。腹鳍退化不存在或仅留存1鳞状硬棘及1退化的软条。臀鰭长,多由分离的短棘组成,有两根刺和56~121个软条。椎骨98~192个。有时会消失。鳔或有或无。
尾鳍小,叉形,或退失消失,使得尾部向后渐尖细。带鱼可根据其尾巴形状特征不同分为叉尾带鱼和细尾带鱼两类。不同属之间存在差异,识别特征如下表:
属名称 | 识别特征 | 图片 |
短尾带鱼属 | 背鳍的硬棘部仅略短于软条部,90~109条 | 黑等鳍叉尾带鱼(Aphanopus carbo) 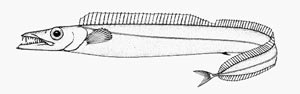
|
棘背带鱼属 | 体长极长;尾鳍呈叉状;腹鳍明显缩小;背鳍116~123条,硬棘与软鳍之间没有缺口;头部有一个突出的矢状嵴。 | 棘背带鱼(Assurger anzac) 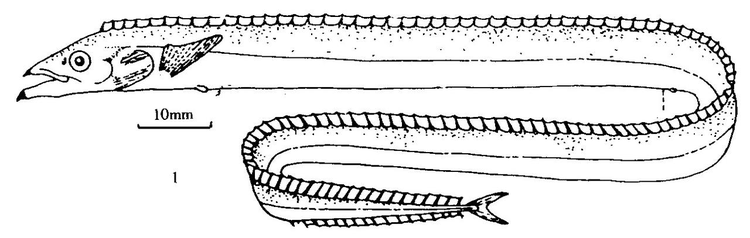
|
深海带鱼属 | 尾鳍小,叉形;腹鳍存在,1鳞状鳍棘和1退化鳍条;背鳍(鳍棘和鳍条)120条以上,鳍棘部为鳍条部的1/2 | 长体深海带鱼(Benthodesmus elongatus) 
|
新月带鱼属 | 下鳃盖大、长,前锯齿状,后缘流苏状;第一肛鳍扩大,匕首状;尾柄椎骨短,长有神经刺(末节椎体除外) | —— |
小带鱼属 | 无腹鳍,成鳞片状突起;下鳃盖骨下缘凸;胸鳍较长,伸达侧线上方;两眼间隔微突;头背缘隆起平缓 | —— |
突额带鱼属 | 头部有突起的矢状嵴;背鳍连续,81~93条;尾鳍小叉状,腹鳍缩小为鳞片状 | —— |
鳞带鱼属 | 头长是总长度的7倍;眼极大,位置接近背线 | 大西洋叉尾带鱼(Lepidopus caudatus) 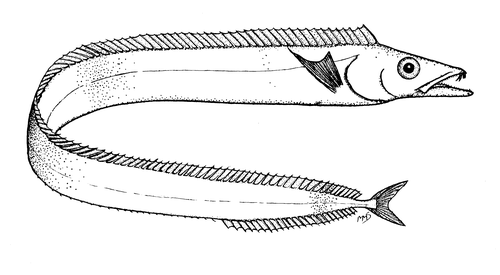
|
沙带鱼属 | 无尾鳍;无腹鳍或腹鳍退化为小鳞片状突起;侧线在胸鳍上方显著下弯;下鳃盖骨下缘内凹 | 沙带鱼(Lepturacanthus savala) 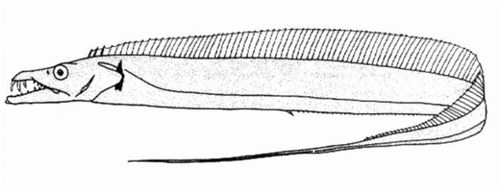
|
狭颅带鱼属 | 胸鳍较短,未伸达侧线;两眼间隔侧扁;头背隆起明显 | 狭颅带鱼(Tentoriceps cristatus) 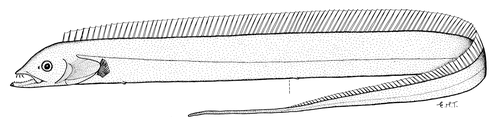
|
带鱼属 | 臀鳍第1鳍棘短,小于瞳孔径;上颌无尖端向前的犬齿 | 珍带鱼(Trichiurus auriga) 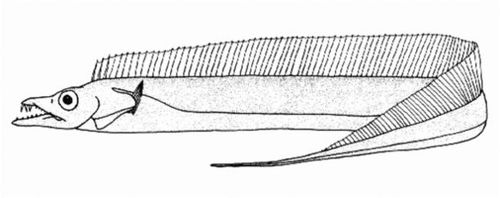
|
分布栖息
分布范围
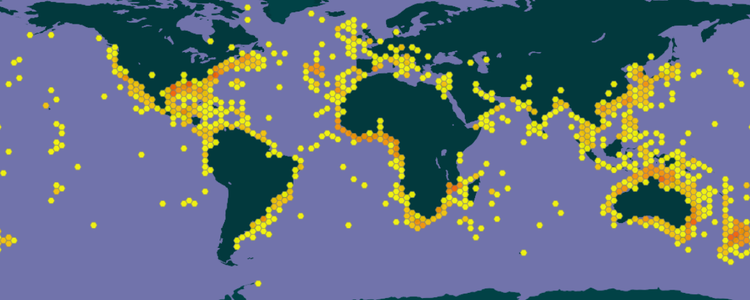
世界分布
带鱼科鱼类广泛分布于大西洋、印度洋、太平洋的热带至温带海域。短尾带鱼属的黑等鳍叉尾带鱼和中等鳍叉尾带鱼(Aphanopus intermedius)是大西洋特有物种,前者分布在北纬71°到北纬22°之间,后者分布在北纬50°到南纬35°之间。印度-澳大利亚群岛有深海带鱼属的四种特有物种组成的集群,少鳍深海带鱼(Benthodesmus oligoradiatus)为印度洋特有种。
中国分布
带鱼科在中国有8属11~12种,主要分布在沿海到开阔海域。中国近海常见的带鱼有带鱼属的日本带鱼(Trichiurus japonicus)、南海带鱼(Trichiurus nanhaiensis)、短带鱼(Trichiurus breviss),沙带鱼属的沙带鱼(Lepturacanthus savala),小带鱼属的小带鱼(Eupleurogrammus muticus)和狭颅带鱼属的狭颅带鱼(Tentoriceps cristatus)。南海带鱼和短带鱼主要分布在中国南海,其他四种在中国分布较广。
栖息环境
带鱼科鱼类是底栖动物,通常栖息于近海大陆架砂泥底质的海域。Aphanopodinae亚科平均栖息深度在330m~1030m的范围,是带鱼中生活海域最深的亚科;Lepidopodinae亚科生活海域比前者稍浅,最深范围在150m~400m;Trichiurinae亚科是带鱼中生存海域最浅的亚科。

生活习性
觅食行为
带鱼科鱼类是广食性、凶猛鱼类。幼鱼主要以小型浮游甲壳类动物为食,成鱼主要以鱼类为食,偶尔吃甲壳类和头足类动物。例如日本带鱼在黄海、渤海,其摄食对象包括虾、乌贼、鱼类等。春季产卵期,其摄食量较少,8-12月,摄食较强盛,昼夜变化不明显。日本带鱼还有食性转换的特点,肛长在20cm以下的日本带鱼,以糠虾类、磷虾类为主食;20cm以上的个体,则以摄食鱼类、甲壳类、头足类为主。不同区域的带鱼食物存在差异。例如黄海的幼鱼栖息在河口时,以糠虾类、毛虾类和小鱼为主要饵料;在浅水区时,则以毛虾类、脊腹褐虾、青鳞沙丁鱼、黄鲫、枪乌贼类等为主要饵料。
带鱼科鱼类的侧线大多位于较低的位置,这是为了适应在海底附近探测外界刺激。它们的牙齿、舌头和鼻器官适应了底栖动物习惯。又尖又长的牙齿用于在伏击捕食时抓住猎物,细圆的舌头不适应在游泳时把水从鳃上引过去,只有一个可以通过水鼻孔,既可以作为流入孔也可以作为流出孔。
节律行为
带鱼科鱼类属于洄游性鱼类,具有生殖洄游习性。在东北亚,该科鱼类主要从北海道南部洄游到日本周围的东海沿岸水域、朝鲜半岛、黄渤海。中国的带鱼科鱼类可以分为南、北两大类,北方带鱼在黄海南部越冬,春天游向渤海,形成春季鱼汛,秋天结群返回越冬地形成秋季鱼汛,南方带鱼每年沿东海西部边缘随季节不同作南北向移动,春季向北作生殖洄游,冬季向南作越冬洄游,故东海带鱼(Trichiurus haumela)有春汛和冬汛之分。
带鱼科成鱼和幼鱼的具有相反的昼夜垂直摄食洄游习性。大型成鱼通常白天在水面附近进食,晚上迁移到底部。白天,幼鱼和小型成鱼在海底100m处成群活动,夜间在地表附近形成松散的摄食集群。
生长繁殖
亲鱼产卵
带鱼科鱼类全年在温暖水域产卵,产卵类型属于全年多次产卵,分批产卵,从而导致带鱼资源常年幼鱼比例较高的现象。同时不同海域带鱼性成熟产卵的时间会有一定的差异,但产卵盛期基本是在夏季,个别海域带鱼在春季和秋季也会出现产卵高峰。例如,南海日本带鱼产卵高峰期从3月份开始,至夏季末期;而在东海,日本带鱼的产卵盛期会比南海稍晚,从5月开始至秋季中期。
带鱼科鱼类是洄游性鱼类,生殖期间会游往近岸或浅海区产卵,日本带鱼每年在产卵期开始产卵洄游,在近河口附近浅海形成产卵群,卵浮性,产卵后成鱼会游向附近渔场索饵,索饵期多在夏、秋季,秋汛渔期过后返回越冬场。黄海、渤海的带鱼种的产卵场位于黄海沿岸各湾和渤海的莱州湾、渤海湾、辽东湾,多在水深20m左右,底层水温14~19℃,盐度27.0~31.0的河口混合水区域。日本带鱼产卵时间很长,黄海、渤海群系的带鱼约为两个月。
带鱼科鱼类是具有浮游卵的硬骨鱼类,卵和幼鱼浮游性,生活在水域上层。其在有利的物理条件下同步产卵,如运输、水平平流、水柱稳定性和沿岸上升流,增加了孵化仔鱼获得食物和避免分散的机会,从而提高了存活率。例如在巴西东南部海域的沿海岛屿附近发现了大量日本带鱼的卵和仔鱼,当地的上升流和洋流可能会提高产量。在表层水温超过21℃的大陆架和大陆坡上发现幼鱼。河口的仔鱼(5~8mm)可在8月份被发现,孵化时间较晚。带鱼可产下多达5196个卵,卵的最大直径为1.1mm。
发育成熟
带鱼科鱼类的雌雄比例通常为1:1或雌性比例较高。带鱼在小群体中的雄性比例较高,而雌性比例则随着体型的增大而增加。在东海,雌性个体比雄性个体更长、更重。由于雌雄带鱼的生长速度和寿命的不同,雌性在2龄以后的生长速度比雄性快,雌性的寿命比雄性长4年以上,因而产生了雌雄差异。幼鱼和亚成鱼出现在沿海水域,成鱼出现在沿海水域到大陆架边缘。
带鱼科鱼类的体长与体重的关系不仅受物种生理因素的影响,还受海域环境条件和性别的影响。例如在黄海、渤海和东海的日本带鱼种群之间的增长存在显著差异。黄海海州湾带鱼雌鱼,第一次性成熟年龄为1~4龄,多数雌鱼性成熟年龄为3龄,雄鱼为2龄。20世纪60年代到80年代,东海带鱼的年龄范围为1~8年,到90年代下降到1~4年,以1~2年龄为主。20世纪90年代,台湾海峡南海带鱼的年龄范围为1~7年,主要集中在3~4年(75%)。
人工养殖
带鱼科鱼类是深海鱼类,有着“离水即死”的“魔咒”,因为当它被捕捞离水后,外界压力突然变低,体内会发生致命变化导致死亡。人为改变压强的成本太高。且带鱼是洄游鱼类,人类无法提供其所需的洄游环境,所以带鱼人工养殖一直是世界性难题。此外,带鱼体表没有大的鳞片,容易受外伤,带鱼生性凶猛,同类之间互相残杀,这些因素都给人工养殖带来了困难。2017年,中国浙江省舟山水产研究所首次成功养成了300多尾东海带鱼,实现人工养殖带鱼技术突破。
物种保护
种群现状
带鱼科鱼类的捕捞对世界渔业做出了重大贡献。全球范围内,海洋渔业资源普遍衰退,但带鱼的产量却一直保持高产且持续增加。据世界粮食与农业组织(FAO)统计,2016年全球带鱼产量约为128万吨,在世界海洋捕捞单种产量中排第13名,2005年-2016年世界带鱼平均年产量约为131万吨,其中,约80%的产量来着中国海洋捕捞。2021年,世界粮食与农业组织报告的三种带鱼(黑等鳍叉尾带鱼、大西洋叉尾带鱼、日本带鱼)全球总捕捞量超113万吨。由于渔业发展,中国海域常见带鱼科物种的种群数量呈现出小型化的趋势。
威胁因素
带鱼科鱼类的致危因素主要是栖息地破坏和过度捕捞。一方面,由于经济发展和气候变化,海洋污染使带鱼的栖息环境遭到破坏。例如东海海域污染,缺氧范围扩大,藻华频繁,肉食性带鱼面临食物来源严重缺乏的问题。日本带鱼在戈亚尼亚河河口受到甘蔗农工业及定居点和村庄污水中汞的生物累积影响。另一方面,过度捕捞打破了种群的自然补偿机制,导致种群无法有效补充,资源减少,产量和个体小型化。如黑等鳍叉尾带鱼在其分布区域内被大量捕捞,其小型个体和未成熟幼鱼占渔获量主要部分。中等鳍叉尾带鱼与由于前者地理界限不清,形态相似,也常被商业捕捞。在地中海的西西里海峡,大西洋叉尾带鱼被过度捕捞。部分种群经常作为副渔获物出现在底拖网渔业中,如日本带鱼。除此之外,有的物种的食物利用能力弱、环境适应能力差、竞争能力弱。使其难以在密集捕捞和气候变化的压力下生存。
保护级别
2004年,有9种带鱼被列入《中国物种红色名录》,其中2种濒危,分别是珠带鱼(Trichiurus margarites)和琼带鱼(Trichiuris minor);2种易危,分别是叉尾带鱼(Benthodesmus tenuis)和狭颅带鱼;3种无危,分别是小带鱼、沙带鱼、日本带鱼;2种数据缺乏,分别是波氏窄颅带鱼(Evoxymetopon poeyi)和条纹窄颅带鱼(Evoxymetopon taeniatus)。
截至2023年,有11种带鱼科鱼类列入《世界自然保护联盟(IUCN)濒危物种红色名录》,其中9种无危(LC),分别是米氏等鳍叉尾带鱼(Aphanopus mikhailini)、中等鳍叉尾带鱼、黑等鳍叉尾带鱼、西蒙深海带鱼(Benthodesmus simonyi)、叉尾带鱼、条纹窄颅带鱼、高额叉尾带鱼(Lepidopus altifrons)、西非叉尾带鱼(Lepidopus dubius)、高鳍带鱼;2种数据缺乏(DD),分别是大西洋叉尾带鱼和潘氏沙带鱼(Lepturacanthus pantului)。
保护措施
主要是限制渔获量和禁止非法捕捞。随着1994年《联合国海洋法公约》和1999年韩国与日本的渔业协定生效,韩国的带鱼科鱼类渔获量持续下降。2012年,国际海洋考察理事会(ICES)建议禁止渔业扩张,减少渔获量,并表明渔业是可持续的。欧洲联盟(EU)对欧盟船只在欧盟以及国际海域捕捞带鱼的总可捕量(TAC)进行管理限制,并在2003年建立许可证制度。针对黑等鳍叉尾带鱼,2015年,欧盟规定的总可捕量为10499吨;2015年、2016年,葡萄牙船队对该物种的渔获量限额分别为3141吨和2827吨(包括中等鳍叉尾带鱼);ICES建议该物种的2017年和2018年的渔获量都不应超过5894吨。黑等鳍叉尾带鱼分布某些区域实施了一些保护措施,即禁止使用有害渔具,例如刺网、拖网和200m以下的底拖网。
另外,还设立保护区保护带鱼的幼鱼繁殖成长,国务院发布了《关于设立幼鱼保护区的决定》,决定从1981年4月22日起,在黄海设立带鱼幼鱼保护区。在保护区内每年8~10月禁止机动底拖网渔船进入生产。农业部发布了《关于国务院批准设立东海产卵带鱼保护区的通知》,决定于1989年起,在中国东海区设立产卵带鱼自然保护区。在保护区内,每年5月1日至6月30日禁止拖网渔船及其他以捕捞产卵带鱼为主的作业渔船进入该保护区内生产。
主要价值
经济价值
带鱼科鱼类的部分种群有重要经济价值,如带鱼属和鳞带鱼属,而有的种群经济意义不大,如深海带鱼属。带鱼科鱼类是中国沿海产量最高的经济鱼类,是中国四大渔业之首,其中带鱼属的日本带鱼渔获量最高,在中国海洋渔业生产中具有重要地位,对渔业生产起着举足轻重的作用。70年代年产量一般在50万吨左右,90年代上升到110多万吨,后来产量不断下降,近几年经过禁渔和开展保护渔业资源方面的宣传教育,比较好地控制了过度捕捞,使带鱼生产保持在一个相对稳定的水平上。
鳞带鱼属的大西洋叉尾带鱼被底层和中上层拖网渔船和延绳钓渔业捕捞,平均上岸量最大的国家是葡萄牙,其中亚速尔群岛贡献了主要的渔获量。从1950年到2019年,开发利用大西洋叉尾带鱼的国家有葡萄牙、南非、新西兰、意大利和西班牙。在亚速尔群岛,大西洋叉尾带鱼是被确定为监测和评估重点的22种鱼类之一。就经济价值而言,该群岛总上岸价值排名第11位,平均每年价值50万欧元。大西洋叉尾带鱼比其他重要的深海经济鱼类长得更大更快,这表明它可能是可持续的渔业资源。黑等鳍叉尾带鱼也是马拉徳群岛和亚速尔群岛的延绳钓渔业捕捞物种,在不列颠群岛西部和西北大西洋的拖网渔业中作为副渔获物被捕捞。在南方的渔获物可能还包括中等鳍叉尾带鱼。
营养价值
带鱼科鱼类肉嫩体肥,味道鲜美,具有很高的营养价值。带鱼肉水分含量74.1%~77.1%,蛋白质16.3%~18.1%,脂肪3.8%~7.4%,灰分1.1%。其脂肪含量高于一般鱼类,且多为不饱和脂肪酸,具有降低胆固醇的作用。对病后体虚、产后乳汁不足和外伤出血等症具有一定的补益作用。中医认为它能和中开胃、暖胃补虚,还有润泽肌肤、美容的功效,不过患有疮、疥的人还是少食为宜。
带鱼肌肉中含有丰富的游离氨基酸,这些氨基酸使带鱼呈现出鲜美的风味。可以晒干、腌制、冷冻、油炸食用,鱼肉可以制作鱼松,内脏可制成鱼粉。带鱼鳞中含有20%~25%的油脂、蛋白质及无机盐,经过酸化可以制成抗恶性肿瘤的硫鸟嘌呤。
代表物种
日本带鱼、黑等鳍叉尾带鱼、大西洋叉尾带鱼具有重要经济意义,棘背带鱼为该科最大种,沙带鱼为带鱼科体型较小种,故将这几个物种作为代表物种进行介绍:
日本带鱼

带鱼属的带鱼(学名:Trichiurus japonicus,同种异名:Trichiurus lepturus、Trichiurus haumela)又称日本带鱼、白带鱼、裙带、白鱼等。体长最大可达234cm。喜煦光,厌强光。摄食习性具季节性,秋季摄食强度最高,春季次之,夏季较低,冬季最低。白天摄食强度高于黑夜。具有渔业经济价值。带鱼肉可以鲜销或制成咸干品。带鱼表皮银膜经提取可制成药用咖啡因。鱼精可提取制精氨酸和核苷酸。
沙带鱼

沙带鱼属的沙带鱼(学名:Lepturacanthus savala,同种异名:Trichiurus armatus、Trichiurus savala)又称珠带、黄带。性贪食,游泳迅速。体型较其他带鱼小,产量少。一般体长为35~50cm,最大可达100cm。分布于印度——西太平洋,中国产于南海、台湾海峡。
黑等鳍叉尾带鱼

短尾带鱼属的黑等鳍叉尾带鱼(学名:Aphanopus carbo)。幼鱼的腹鳍只有一根刺,成年后完全退化。体色为铜黑色,带有彩虹色。最大体长可达145cm,80cm~85cm时性成熟。成熟的个体会向马德拉群岛和加那利群岛附近的产卵地和育儿地进行水平迁移。商业渔获量达1000吨。
棘背带鱼

棘背带鱼属的棘背带鱼(学名:Assurger anzac)。最大体长可达225cm。分布较广,在大西洋的波多黎各岛和乌拉圭等海域,印度洋的澳大利亚西岸海域,太平洋的新几内亚、日本南部、中途岛及加利福尼亚等沿海150m~400m的中深海域均有分布。体无鳞,除两颌前端的皮质突起为黑褐色外,全体银白色。
大西洋叉尾带鱼

鳞带鱼属的大西洋叉尾带鱼(学名:Lepidopus caudatus)。整体均匀银色。腹鳍极小。最大体长可达210cm。在东大西洋、西地中海、南印度洋以及澳大利亚和新西兰附近的渔业中都具有重要价值。通常分布在100m~250m的沙质和泥质海底,在澳大利亚超过300m,在爱奥尼亚海东部的深度范围为333m~620m。夜间迁徙至中层水域形成集群。偶尔出现在近岸的上升水流中。