简介
细尾獴属于非洲西南部干旱地区特有种,在当地很常见,于2015年被IUCN红色名录评定为无危(LC)。
分类和演化
分类
细尾獴为细尾獴属(Suricata)下的唯一一种物种。1776年,德国博物学家施雷贝尔(Johann Christian Daniel von Schreber)依据一件从好望角发现的标本首次描述了该物种,但由于施雷贝尔误以为细尾獴是一种灵猫,而导致它被施雷贝尔错误地归类到了灵猫属(Viverra)中并定名为Viverra suricatta。后来,法国动物学家德马雷(Anselme Gaëtan Desmarest)意识到细尾獴并不应该属于灵猫属,他在1804年首次建立了细尾獴属(Suricata),并依据另一件来自好望角的标本为新种“开普细尾獴”(Suricata capensis)命名。英国动物学家托马斯(Oldfield Thomas)和施万(Harold Schwann)首次提出了该种正确学名应该为Suricata suricatta,并指出开普细尾獴是无效名,它只是细尾獴的异名同物。

目前,已经确认世界上有三个细尾獴亚种,但是这三个亚种的有效性未经分子生物学检验。
演化和系统发育
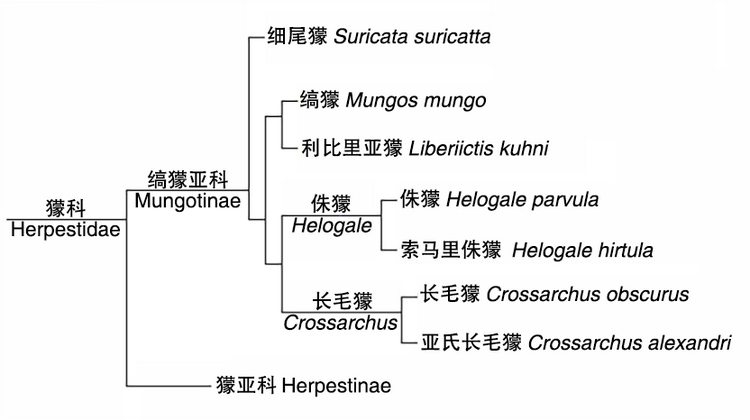
分子生物学研究显示,獴科(Herpestidae)起源于距今约2230万年前。到大约1460万年前,该科分化为两个演化支,当时由于森林栖息地减少,一部分早期獴类被迫来到草原上生活,发展出社会性行为,最终演化为今天的群居獴类——缟獴亚科(Mungotinae);而留在森林里的那部分獴类仍很大程度上保留了独居的生活习性,演化成为今天的獴亚科(Herpestinae)。细尾獴属于缟獴亚科成员,该亚科成员还包括长毛獴(Crossarchus)、侏獴(Helogale)、利比里亚獴(Liberiictis)以及缟獴(Mungos)等。细尾獴是该亚科中最早分化出来的一种,它早在1250万年前就与该亚科其他成员分离了。
由于獴科的起源和演化长期发生在旧大陆的热带地区,而热带地区的环境条件不利于化石形成,因此细尾獴乃至整个獴科的化石材料都很缺乏,其演化史在很大程度上仍是未知的。少量的细尾獴化石材料曾出土于南非德兰士瓦的Hearths洞穴,以及萨尔达尼亚附近的Hoedjiesbaai化石点,年代为更新世(距今260~1万年前)。在南非埃兰兹方丹化石点发现的大细尾獴(Suricata major),其生存年代为距今约180万年前,被认为是细尾獴类的早期成员。大细尾獴的解剖结构介于现代细尾獴和缟獴(Mungos mungo)之间,这也侧面印证了细尾獴和缟獴确实存在较近的亲缘关系。但是,目前归于该种的化石材料形态差异很大,可能并不属于一个单一物种,这些材料的归属尚有待于进一步鉴定。
形态特征
细尾獴是一种小型獴类,在獴科中仅比侏獴略大;体长24.5~29cm,尾长19~24cm,后足长5.9~6.8cm,耳长1.6~2.2cm,雄性平均重731g(范围是626~797g),雌性平均重720g(范围是620~969g)。脸颊呈锥形,前额部圆,耳呈新月形。身体纤细,四肢细长;尾细长,往尖端渐细,不似多数獴类那样尾毛蓬松,细尾獴正是因此而得名。细尾獴常采取一种两足直立的姿势,此时长而有力的尾巴可以辅助支撑身体。

毛色通常为浅灰色、黄褐色或棕褐色,存在明显的地理变异,南部的个体毛色较深,而生活在其分布区西部和西北部干旱地带的个体毛色较浅。体毛柔软且紧贴身体。毛分两层,外层刚毛比较长,在身体大部分区域长15~20mm,在体侧可长达30~40mm,有些个体的刚毛基部呈浅色,中部有两个深色的环,尖端为银白色或黑色。里层绒毛稀疏,通常为深棕色,有助于加快热量传播。头部大部分为近白色,两眼周围有独特黑色眼斑,外形和功能均似太阳眼镜;鼻子为褐色,上面覆盖有短毛;耳朵为黑色。

身体背面有较暗淡的斑点,从肩部一直延伸到尾部,这些斑点有时会融合成横纹,在腰部最多见。身体腹面为米色或黄色。


细尾獴的身体构造非常适于挖掘洞穴和在洞穴中穿行,它的足部狭窄,前后足均仅有四趾,拇趾退化,其余四趾紧靠在一起。爪子高度发达,在整个猫形亚目中都是首屈一指。体侧的长刚毛有一定的触觉功能。耳比多数獴类要小得多,在挖洞的时候可以闭合,以防泥土溅入。

细尾獴头骨的最大特点是眼眶很大,超过颅全长的20% 。头骨构造轻盈,较高,整体形状呈圆形。头骨上没有明显的矢状脊,人字脊亦不发达,颧弓细小,冠状突高度中等,这些附着咀嚼肌的部位均不发达,说明细尾獴咬合力较小。共有36颗牙,齿式为3.1.3.2/3.1.3.2(上颌每侧各有3颗门齿、1颗犬齿、3颗前臼齿和2颗臼齿,下颌每侧各有3颗门齿、1颗犬齿、3颗前臼齿和2颗臼齿),门齿列呈弧形,犬齿相对细小,裂齿较退化,臼齿宽大且齿尖锋利,其牙齿结构适于碾碎昆虫,而不适合捕杀较大的猎物。

分布信息
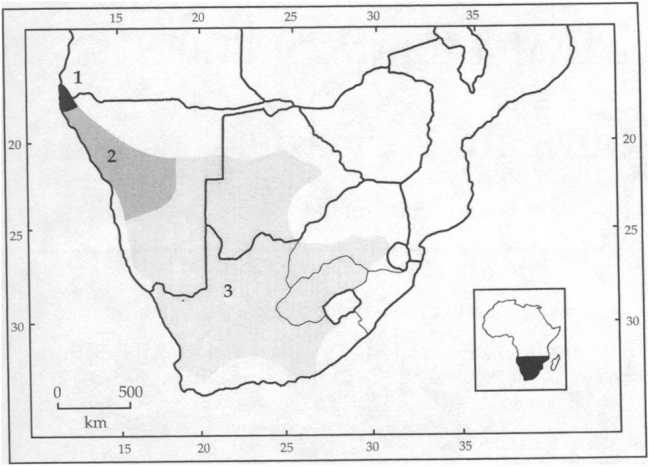
地理分布
细尾獴在南非地区西半部广泛分布。在纳米比亚,该种见于西部和南部的大部分地区,向西北延伸到安哥拉西南角。该种亦出现在博茨瓦纳西南大部,北至21°S,东至马卡迪卡迪盐沼盆地(Makgadikgadi Pan)。该种在南非共和国大部分地区都有分布,唯独不见于该国东北角、最东端和南部沿海地带;且在该国西半部比东半部更常见。有学者认为该种在莱索托的低海拔地区亦有分布,但迄今未发现可信记录。
栖息地
细尾獴栖身于干旱、开阔的生境,它的偏好生境特点是草短,灌木和乔木稀疏;其栖息环境的干旱、开阔程度为獴科之最。栖息地类型包括热带、亚热带干旱的草原和灌木丛,以及半荒漠地区,但不会出现在真正的沙漠,在森林和山区也没有该种分布。
细尾獴的生境选择很大程度上受限于土壤类型,由于高度依赖洞穴生活,它多生活在以沙质土为主的地区;不过,在南非共和国很多地区,尽管土质不是那么松软、易碎,但依然有细尾獴出现。
生活习性
觅食习性
细尾獴以无脊椎动物为主要食物,亦食小型脊椎动物以及鸟和爬行类的卵,有时也吃一些植物性食物。其中昆虫是细尾獴最主要的食物,在其食谱中占比超过八成;食用最多的昆虫包括5类:鞘翅目、鳞翅目、等翅目、直翅目和双翅目。其食谱因季节而异,冬季主要取食鞘翅目和鳞翅目,夏季则以鞘翅目和等翅目为主食。除昆虫外,蛛形类、多足类等其他节肢动物在其食谱中也占有重要地位。细尾獴可捕食蝎子、马陆等毒虫,对它们的毒液有抵抗力

脊椎动物在细尾獴食谱中的占比很小,通常小于6%。根据野外观察,细尾獴取食过的小型脊椎动物包括小型鸟类(南方蚁鵖)、小型爬行类和小型两栖类。曾有报道称细尾獴能捕杀小型鼠类,但在野外从未有过记录。不过,在动物园里曾有人观察到,一只细尾獴未经任何训练即一口咬死了一只小型哺乳动物。
在野外细尾獴基本是虫食性动物,很少取食植物。只有当缺水的时候才会啃食瓜果、块根和块茎,以此来补充水分。但在动物园中,它会变成一种杂食性动物,对人们喂食的水果、蔬菜来者不拒。

细尾獴每天花5~8个小时觅食。虽是群居动物,但它与其他群居獴类一样通常独自觅食。不过,群体成员会彼此在对方的视野内觅食,并以叫声保持联系。细尾獴不会采取“守株待兔”式的机会主义觅食,而是非常积极地搜寻食物。不同个体走不同的觅食路线,沿途翻开石头、挖开草根,彻底搜寻所经之处的所有猎物。每天变换觅食路线,每搜寻一处后一周之内不会返回,为食物恢复留足时间。细尾獴嗅觉发达,而听觉相对退化,故通常以气味定位猎物,用发达的前爪将猎物从土里挖出来。成年细尾獴会乐意与群内的幼仔分享食物。细尾獴通常不会追逐猎物,偶尔追逐小蜥蜴亦不会超过几米远。冬季食物摄入量通常低于夏季。
社会行为
细尾獴是一种高度社会化的动物,生活在2~30只个体的群体中,平均每群有15只个体,最多达49只。每个群体的家域面积平均为5km² ,大者可达15km² 。家域内包含很多巢穴,这些巢穴彼此间相隔50~100m。它们会因食物缺乏、洪水或天敌的威胁而迁徙,轮流使用家域内的多个巢穴 。群体内彼此友好,有时会发生仪式化的打斗,但不会造成伤害。群体之间则充满敌意,彼此相遇会发生严重打斗,甚至造成死亡,说明这种动物可能存在较强的领域性。

群体成员共同担负照顾幼仔和警戒天敌的职责。群内有明显的社会等级,通常体型最大的雌性地位最高。一般来说,只有少数优势个体有繁殖的权利,它们会杀死次等个体的幼仔,令次等个体只能无私地照顾它们的幼仔。如果次等雌性能成功养育自己的幼仔,它有时也会杀死优势雌性的幼仔,以提高自己后代的地位。
为求得繁殖机会,雄性的次等个体完全长成后,会主动离开原生群体,积极向外扩散。它们刚与新群接触的时候会遭到新伙伴的排斥,往往要花几天时间才能得到接纳。雌性个体则通常会选择留在原生群体中,随着它们年龄增长,地位逐渐提高,将有可能获得繁殖的权利。也有些雌性个体会选择扩散,但它们多倾向于自己招募流浪雄性,新建一个群,或者加盟没有繁殖雌性的群体,而不太可能得到本就有雌性的群体接纳。
穴居行为
细尾獴营穴居生活,它那发达的爪子非常适合挖洞。当建造或翻修巢穴时,一群细尾獴排成一列,前面的个体用前爪捣碎泥土,推到身后,再用后爪传递给后面的个体,直至将土推出洞外。不过,细尾獴更喜欢占用现成的洞穴,包括天然形成的石洞以及其他小型哺乳动物建好的巢穴。它常与南非地松鼠(Xerus inauris)、黄獴(Cynictis penicillata)混居,学者发现细尾獴85%的巢穴均是与其他动物混居的,混居个体是不混居个体的6倍。
细尾獴巢穴内的构造可能很简单,也可能复杂如迷宫。通常,巢穴占地范围直径约5m,有大概15个入口;穴内通道常有两到三层,直径约15cm,与地面成40°左右的夹角向下倾斜;每个“居室”30cm高,15~45cm长,里面没有枯草衬垫。1940年有学者报道了一个极为复杂的细尾獴巢穴,占地面积为25×32m,有多达90个入口。当天敌来袭时,更多的入口能给细尾獴带来更多的逃生机会。

巢穴内温差很小,当地年度温差可达43℃(夏季21~39℃,冬季-4~26℃),而巢穴内年度温差13.2℃,昼夜温差不到1℃(夏季22.6~23.2℃,冬季10.0~10.8℃)。巢穴为细尾獴创造了一个非常宜居的微环境,有效减小了它的体温调节成本,对这种体毛稀疏的小型恒温动物来说至关重要,因此细尾獴会在巢穴内度过大量时间。它是严格的昼行性动物,每天清晨出穴,以两足直立的姿势聚集在洞口,梳理毛并晒太阳;天黑前即返回穴内,聚在一起睡觉;夏季正午还要躲进穴内避暑。据统计,在冬季,细尾獴每天在巢穴内度过一半的时间,而夏季则要度过大约三分之二的时间,遇到严寒和阴雨天气会在里面呆得更久,甚至一天不出巢穴。
生长繁殖
细尾獴的交配季节通常与雨季同期,例如在卡拉哈里南部是10月到1月;在多雨年份,交配季节也会有所延长。多数幼仔在1~3月出生,但其他月份亦有幼仔出生记录。在降水量特别低的年份,细尾獴可能全年不繁殖。通常只有优势雌性有繁殖的权利,不过,在个别降水量高、食物丰富的年份,次等雌性也有机会繁殖。雌性一岁即性成熟,动物园里有14月龄雌性繁殖的记录,但在野外通常要到3岁以后才有繁殖机会。优势雌性每年产1~3胎幼仔,平均1.9胎;每胎产3~7只幼仔;在降水量较高的年份产仔数更多。次等雌性产仔数比优势雌性少。
雌性细尾獴妊娠期约60~70天,幼仔约19日龄开始走出巢穴自由活动,26日龄开始跟随群体一起觅食,49~63日龄断奶。新生仔重约100g,6周龄增重到179g,3月龄重394g,6月龄达到529g,1岁时达到成体体重的大约90%。
群体合作育幼,未参与繁殖的次等个体会主动充当照顾幼仔的助手。当幼仔还小不能离开巢穴的时候,助手要在母亲外出觅食时充当“保姆”,或者为“保姆”提供食物。“保姆”的工作不仅包括守卫幼仔,还包括用自己的身体给幼仔取暖,防止发生幼仔体温过低导致死亡的情况。大约四分之一的群体存在异亲哺乳现象,即群体内未繁殖的雌性在优势雌性分娩后,发生了自发泌乳,并参与为幼仔哺乳。当幼仔可以跟着成体一起外出觅食以后,助手负责看护幼仔,并给幼仔喂食。助手对幼仔的成长至关重要,有研究表明,参与照顾幼仔的助手数量不仅会影响幼仔的体重增长率,还会影响到幼仔的首次繁殖年龄,甚至它日后成为优势个体的可能性。
幼仔跟随群体外出觅食时,会与成体保持2m内的距离,并持续发出一种类似于鸟鸣的叫声,向成体索求食物。而当成体发现食物,会发出一种音调更高的叫声,招呼幼仔前来觅食。研究发现,幼仔越饿,叫声越响亮,持续时间也更长,成体以此来判断幼仔的饥饿程度,优先给更饥饿的幼仔喂食。随着幼仔成长,成体会让幼仔与活体猎物接触,以此磨砺幼仔的觅食能力。例如,有学者曾观察到,一只成年细尾獴给幼仔示范吃蝎子之前要先去掉它的毒刺。幼仔约90日龄开始独立觅食,它到100~120日龄时无法再发出类似于鸟鸣的求食叫声,此时成体也会停止给它提供食物。
助手提供给幼仔的食物量多少不定,有时候它们会把自己发现的食物近一半都喂给了幼仔,而有时候只喂给幼仔3%,平均值为26%。由于助手要优先保证自己的生存,当它发现的食物较少的时候,它提供给幼仔的比例也较低。喂食中还存在性别偏见现象,雌性助手主要给雌性幼仔喂食,而雄性助手则平等地给两性幼仔喂食。这可能是因为,雌性通常一生都留在原生群体中,而对原生群体来说,雌性幼仔的存活更有价值。
细尾獴群体中雄雌性别约为1:1,幼体占比约为30%。幼仔存活率取决于群体大小、气温和降水量。幼仔存活需要适度的温度和降水;在高温、干旱的年份,幼仔生长缓慢,存活率也下降;而3~5月龄个体最主要死亡原因即是因获取热量不足而被冻死;洪水亦会导致幼仔被淹死在巢里。此外,小幼仔时常死于同类成年个体的“杀婴行为”。而对超过5月龄的大幼仔和成体来说,天敌捕食是最主要的死因。成体死亡率约32~68%,取决于天敌密度;而幼体死亡率高达76~80%。在野外寿命有5~15岁,而在饲养状态下有活到20.6岁的记录。

种间关系
细尾獴最大的天敌是猛禽,包括短尾雕、猛雕、茶色雕和淡色歌鹰。它对猛禽非常警觉,当群体多数成员忙于觅食时,总有一些个体负责像哨兵一样“站岗”,一旦发现猛禽来袭就持续发出一种高音调的警戒叫声,提醒其他个体逃跑。这种动物不擅奔跑,亦不擅攀爬,最佳御敌策略就是尽快躲进洞里。除猛禽外,一些猫科和犬科动物都有可能危害细尾獴。可能威胁细尾獴的犬科动物包括黑背胡狼、南非狐和蝠耳犬。而在猫科动物中,大至狮子、小至非洲野猫都有捕食细尾獴的记录。
细尾獴常与其他穴居动物混居,共用一穴,最常见的是与南非地松鼠(Xerus inauris)、黄獴(Cynictis penicillata)混居。考虑到这几种穴居动物均有季节性迁徙的习性,混居可提高巢穴利用率,节省动物的时间和精力,对参与的所有动物都有好处。细尾獴与南非地松鼠在食物和空间上不存在竞争,因此它们可以毫无敌意地共处一穴。细尾獴与黄獴之间的竞争程度也很低,因为黄獴的分布远比细尾獴广泛,而且黄獴捕杀鼠类、鸟类和蛙类等小型脊椎动物比细尾獴容易得多。有记录与细尾獴混居的小型动物有十几种。


物种保护
细尾獴当前没有面临重大威胁,种群数量保持稳定,于2015年被世界自然保护联盟(IUCN)红色名录评定为无危物种。其种群密度在不同地区差异很大,受降水量和天敌的影响。在南非共和国的卡拉哈迪跨界国家公园,1994年4月细尾獴的密度为1只/km²,由于降水量减少,到1995年5月密度即下降到仅有0.32只/km²。主要的长期研究项目为克鲁顿-布罗克(Tim Clutton-Brock)发起的“卡拉哈里细尾獴项目”,该项目旨在研究该种的社会行为。
与人类的关系
细尾獴不会危害家禽,相反会捕食大量农业害虫,因此被农场主认为是益兽而加以保护。而且该种在野外挖掘洞穴,具有重要生态价值。主要危害是传播狂犬病。尽管该种患狂犬病而攻击人畜的记录很少,而黄獴才是南非地区最大的狂犬病传染源,但该种有时还是会在当地政府组织的狂犬病控制行动中被一并清除。
由于该种外形可爱,深受人们喜爱,因此频繁出现在电影、书籍等文艺作品中。1987年,BBC野生动物纪录片《细尾獴国度》的上映,使细尾獴的形象开始深入人心。迪士尼动画片《狮子王》中的角色“丁满”原型即是一只细尾獴。英国牛津科学电影公司制作的电视连续剧《细尾獴大宅门》以“卡拉哈里细尾獴项目”研究的一群细尾獴为主角,将肥皂剧的手法和纪录片风格的镜头相结合,向大众介绍了该项目的一些研究成果;该剧于2005年上映,受到广泛好评。
由于细尾獴是非洲小型哺乳动物的代表性物种,世界上有很多家动物园愿意饲养并展出细尾獴。也有人会捕捉野生细尾獴当宠物饲养,这种做法尚未对野生种群产生显著危害,但由于该种攻击性较强,实际上并不适合作为宠物。
注释
[a]中国著名动物学家谭邦杰编著的《哺乳动物分类名录》与中科院动物研究所研究员王海滨译著的《世界陆生食物动物大百科》中均使用了“细尾獴”做为该动物的中文正式学名。“狐獴”一名更多用于文学作品中。












